
| Este artigo tem como objetivo a crítica da doutrina da macroevolução, particularmente sob a forma em que é ela apresentada nos colégios e faculdades. É citado o conhecido livro de Lasker “Antropologia Física” (1), para mostrar como a doutrina é apresentada de fato. Citações de numerosos autores indicam que praticamente todas as hipóteses da doutrina da macroevolução são, no mínimo, questionáveis.
Deve ser entendido que este artigo não tem a intenção de atacar Lasker nem seu livro. Pelo contrário, constitui uma crítica da doutrina que o autor esposa em seu livro. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Introdução A tese que se defende neste artigo é que a doutrina da evolução no nível “macro” não tem apoio científico real. Não se questiona a “microevolução” (exceto, possivelmente, quanto à impropriedade de sua designação), isto é, o tipo de alteração pela qual por exemplo os geneticistas desenvolvem cachorrinhos do tamanho da palma da mão, ou laranjas com maior quantidade de suco. Faz-se objeção à tentativa de extrapolar essas pequenas alterações para supostas alterações macroevolutivas, pelas quais hipoteticamente os peixes se transformaram em gente, ou partículas irracionais aleatórias se transformaram em organismos progressivamente mais complexos.O livro do Professor Lasker “Antropologia Fisica” (1) é um texto popular e largamente adotado. Ele apresenta a doutrina evolucionista atual de maneira bastante precisa. Por essas razões, foi ele escolhido como o meio pelo qual o autor deste artigo apresenta seus próprios pontos de vista contrários.As críticas que seguem, portanto, são dirigidas não ao Professor Lasker, mas à doutrina sobre a qual se baseia o seu livro. Como ficará claro, tais criticas tornam-se possíveis em grande parte devido à disposição dos pensadores evolucionistas em admitir francamente a existência de áreas problemáticas. A tese que se defende neste artigo é que a doutrina da evolução no nível “macro” não tem apoio científico real. Não se questiona a “microevolução” (exceto, possivelmente, quanto à impropriedade de sua designação), isto é, o tipo de alteração pela qual por exemplo os geneticistas desenvolvem cachorrinhos do tamanho da palma da mão, ou laranjas com maior quantidade de suco. Faz-se objeção à tentativa de extrapolar essas pequenas alterações para supostas alterações macroevolutivas, pelas quais hipoteticamente os peixes se transformaram em gente, ou partículas irracionais aleatórias se transformaram em organismos progressivamente mais complexos.O livro do Professor Lasker “Antropologia Fisica” (1) é um texto popular e largamente adotado. Ele apresenta a doutrina evolucionista atual de maneira bastante precisa. Por essas razões, foi ele escolhido como o meio pelo qual o autor deste artigo apresenta seus próprios pontos de vista contrários.As críticas que seguem, portanto, são dirigidas não ao Professor Lasker, mas à doutrina sobre a qual se baseia o seu livro. Como ficará claro, tais criticas tornam-se possíveis em grande parte devido à disposição dos pensadores evolucionistas em admitir francamente a existência de áreas problemáticas.
Obviamente, todos os evolucionistas citados neste artigo permanecem evolucionistas, mesmo admitindo a existência de problemas em seus próprios campos. Às vezes supõem eles que as evidências de outros campos, apesar de tudo, continuam a apoiar a evolução, ou que futuras descobertas de algum modo defenderão a doutrina, apesar dos fatos contrários que hoje se conhecem. Ao se considerar a extensão e a profundidade dos problemas, conclui-se que toda a doutrina é questionada. À medida em que os problemas sejam levantados neste artigo, e essa é a sua principal finalidade, o leitor cuidadoso poderá referir-se às fontes originais para julgar por si mesmo se a doutrina da macroevolução é adequada, ou sequer razoável. Pode-se iniciar, como o Professor Lasker, com a origem da vida. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A Origem da VidaNa última metade do século XVII, Francesco Redi demonstrou que, contrariamente ao que muitos acreditavam, as moscas não se geravam espontaneamente na carne em putrefação. Louis Pasteur demonstrou em 1861 que também as bactérias não se geravam espontaneamente no leite. Não obstante, a geração espontânea continua sendo um ingrediente essencial na doutrina evolucionista.
Quais são as evidências? O Professor Lasker declara: “Tanto quanto saibamos, os compostos orgânicos que ocorrem naturalmente hoje são resultantes de processos vitais; a vida parece sempre depender de vida previamente existente” (página 26). Ressalte-se que, neste contexto, “orgânico” não significa “com vida”, mas simplesmente qualquer composto contendo Carbono. Tendo assim exposto os fatos, Lasker continua a especular nas páginas 26 a 28: “Imagine-se, então, uma época anterior à existência da vida. Mesmo que os compostos orgânicos não estivessem sendo sintetizados nas células, pelo menos não haveria microorganismos que os degradassem, a partir do estado em que existissem. Não havendo nada que obrigasse os compostos a se dissociar, a evolução teria ocasionado compostos cada vez mais complexos, e não mais simples…”. Alguns parágrafos adiante, acrescenta ele: “De qualquer maneira, os constituintes das proteínas e dos ácidos nuclêicos das células vivas foram sintetizados em laboratório, sob as condições existentes na Terra primitiva. Embora ninguém tenha ainda chegado perto da criação de uma célula viva a partir do ar primitivo, não obstante têm-se mostrado plausíveis os passos químicos necessários” (página 32). A “plausibilidade” desses passos deveria ser considerada mais detalhadamente. Primeiramente, não é correto dizer que, na ausência de microorganismos, “nada houvesse que obrigasse os compostos a se dissociar”. Nas experiências citadas por Lasker (realizadas por Miller, Sagan e Khare, Wollin e Ericson, Stephen Sherwood e Oparin) os aminoácidos e outros compostos simples tiveram de ser removidos rapidamente da presença da fonte de energia que os produzia (centelhas elétricas ou luz ultravioleta) porque a taxa de sua destruição sob a sua ação, ou sob a ação de quaisquer outras fontes de energia propostas, excedia de muito a taxa de sua produção (2). Mesmo que se supusesse que esses compostos pudessem ter-se formado, e permanecido na atmosfera durante tempo suficiente para chegar até o oceano, a maioria deles teria sido destruída pela água nas temperaturas comuns. Por exemplo, Miller e Orgel declararam que “as taxas de depurinação do DNA, da hidrólise dos polímeros peptídicos e polinucleotídicos, e da decomposição dos açúcares, são tão grandes, que parece impossível que tais compostos pudessem ter-se acumulado em solução aquosa e pudessem ter sido usados nos primeiros organismos, a menos que a temperatura fosse baixa” (3). Tais temperaturas baixas, entretanto (Miller e Orgel preferem 0 °C ou menos), parecem incompatíveis com a hipótese evolucionista de que a vida se tenha formado logo após a Terra ter-se resfriado, a partir de uma massa fundida, enquanto os oceanos ainda estavam muito quentes.
Parece, entretanto, que mesmo que originalmente não houvesse Oxigênio, a fotólise do vapor d’água atmosférico, sob a ação da luz ultravioleta, teria produzido uma quantidade significativa de Oxigênio bem cedo na história da Terra (4). Além disso, não há evidências de que a atmosfera jamais tenha contido metana (5), que é o gás mais comumente utilizado nas experiências que supostamente têm a ver com a origem da vida. A amônia, que, juntamente com a metana, é necessária para a produção dos compostos, também é sujeita a fotólise rápida, sob a ação da luz ultravioleta, pelo que ter-se-ia reduzido a concentrações muito mais baixas do que as usadas nas experiências (6). Suponha-se, entretanto, a título de argumentação, que os aminoácidos pudessem ter-se formado. Poderiam eles ter-se arranjado por si mesmos em forma de “cadeias” para formarem proteínas? Lasker afirma: “Ao tentar a síntese abiogênica de proteínas, S. W. Fox (1960) conseguiu produzir, a partir de aminoácidos, substâncias que, sob alguns aspectos, assemelham-se a proteínas. Elas são capazes de assumir a forma de pequenas esferas uniformes que podem ser vistas ao microscópio. Em alguns casos o microscópio eletrônico revela diferenciação e características semelhantes às de células, como por exemplo membranas de parede dupla nas gotículas” (página 33). Sidney Fox produziu essas cadeias de aminoácidos pelo aquecimento de uma mistura pura e seca de certos aminoácidos, usualmente durante 6 a 10 horas, a 170-200 °C, ou por uma semana a 120 °C. Em seguida, agitou-a em água quente, removendo o material insolúvel por filtragem. Resfriando a solução, precipitaram-se glóbulos microscópicos, ou “microesferas”, contendo as cadeias de aminoácidos. Fox alega que essas “microesferas proteinóides” são estáveis e exibem uma espécie de evolução pré-biológica. Entretanto, Oparin afirma: “As microesferas de Fox, sendo obtidas termicamente, não apresentam resultados muito promissores, deste ponto de vista (isto é, da evolução até incluir processos metabólicos). Sua estrutura é estática. Isso … cria muitas dificuldades quando se tem de convertê-las em sistemas dinâmicos que possam ser utilizados como modelos da evolução do metabolismo” (7). (Ênfase acrescentada). De fato, as experiências de Fox dependem de um suprimento de aminoácidos puros e secos, necessidade esta que parece ser impossível satisfazer, como foi observado. Se tal suprimento, entretanto, fosse disponível, Fox mostrou que, sob certas condições, seria possível ligar entre si os aminoácidos. Ele sugere, então, que tais condições deveriam ter existido na Terra primitiva, nas bordas dos vulcões. Entretanto, Miller e Orgel criticam essa sugestão, ressaltando que, ao se solidificar, a lava é muito mais quente do que o ar, e acrescentam: “Outra maneira de examinar este problema é perguntar se existem locais na Terra, hoje, com temperaturas adequadas, em que pudéssemos por exemplo semear 10 gramas de uma mistura de aminoácidos, e obter uma colheita significativa de polipeptídeos … Não podemos sequer imaginar um local semelhante” (8). De maior significado é ainda a declaração de Fox de que seus polipeptídeos (cadeias de aminoácidos, mais curtas do que as proteínas) são semelhantes às proteínas na ordem em que as ligações são formadas. O fato de que mesmo as mais simples proteínas exigem uma ordem definida na ligação de seus aminoácidos, e de que é virtualmente nula a probabilidade de que tal ordem ocorra espontaneamente, tem sido fonte de consideráveis desapontamentos aos defensores da geração espontânea. Tanto quanto se saiba, nada há, no que se refere às propriedades fisicas e químicas dos aminoácidos, nucleotídeos (os blocos construtores do DNA), e açúcares, que indique que as polimerizações envolvendo misturas de quaisquer dessas substâncias, produzam algo além de seqüências ou estruturas aleatórias. Essa aleatoriedade foi demonstrada experimentalmente por dois companheiros de Fox, Peter Mora e Gerhard Schramm, na polimerização de açúcares (9), e de nucleotídeos (10). As evidências contrárias relativamente às cadeias de aminoácidos de Fox foram comentadas por Miller e Orgel, que chegaram a acusá-lo de fraude: “O grau de não-aleatoriedade dos polipeptídeos térmicos assim demonstrado é mínimo em comparação com a não-aleatoriedade das proteínas. É um erro, portanto, sugerir que os polipeptídeos térmicos sejam semelhantes às proteínas em sua não-aleatoriedade” (11). Um dos fatos mais misteriosos da Biologia relaciona-se com esse problema de ordem em contraposição à aleatoriedade. Todos os aminoácidos encontrados nas proteínas, com uma única exceção, a glicina, podem existir em duas formas designadas por D e L (dextrógira e levógira). Química e fisicamente elas exibem propriedades idênticas, com exceção de certas propriedades óticas que não pareceriam ter nada a ver com a sua utilização biológica. Biologicamente, porém, a diferença é enorme, pois todas as proteínas produzidas nas células vivas contêm exclusivamente a forma L, ou seja a levógira. Todas as proteínas produzidas artificialmente contêm, entretanto, misturas aleatórias das formas D e L. A substituição de um simples aminoácido em uma proteína, por outro com a forma D, destrói completamente toda a sua atividade biológica (12). Levando-se isso tudo em conta, a alegação de Fox quanto à não-aleatoriedade, parece realmente questionável. À vista dos problemas trazidos á luz pelas experiências mencionadas, a geração espontânea parece cada vez menos plausível. Considerem-se as cifras seguintes, apresentadas por Gerhard Schramm, com relação à probabilidade de formação ao acaso de uma das mais simples formas de vida (alguns chegam mesmo a questionar se ela devesse sequer ser chamada de forma de vida), o vírus do mosaico do fumo: “O ácido ribonuclêico (RNA) do vírus do mosaico do fumo contém 6000 nucleotídeos. A probabilidade de que essa molécula especial resulte da combinação aleatória de quatro nucleotídeos é (1/4)6000 = 10 –2000. Considerando que todo o universo contém o número estimado de 1080 prótons, é praticamente impossível obter-se esse ácido ribonuclêico no período de 109 anos, a idade estimada da Terra, mesmo que toda a Terra consistisse de uma mistura de nucleotídeos em reação” (13) . Schramm e outros defensores da geração espontânea tentam libertar-se desse dilema mediante alguma espécie de “seleção natural” para as moléculas pré-biológicas. Entretanto, como essas moléculas não são auto-replicativas, isso constitui uma contradição (14). Peter Mora é mais objetivo. Comentando a respeito da prática usual dos evolucionistas de “apelar para o infinito”, afirma ele: “Creio que desenvolvemos essa prática para evitar enfrentar a conclusão de que a probabilidade de um estado auto-replicativo é zero. … Ao se ter de invocar, para propósitos práticos, o conceito de tempo e matéria infinitos, o conceito de probabilidade é anulado. Mediante tal lógica pode-se provar qualquer coisa, como por exemplo, que qualquer coisa se auto-reproduza, exatamente e imensuravelmente, independentemente de sua complexidade” (15). Mora sugere, com propriedade, que “a ilusão de que o problema possa ser explicado com o conhecimento existente” constitui uma “atitude mental perigosa”, e que deveríamos “mesmo ousar perguntar se não há nos seres vivos algo especial que não pode ser tratado pela Física, tal qual a conhecemos, mas que deva ainda ser submetido a comprovação ou refutação” (16). Mutações Mesmo que o problema da geração espontânea pudesse ser superado, como uma espécie de organismo se transformaria em outra, diferente, mais complexa? O Professor Lasker admite que “A única maneira pela qual surgem possibilidades totalmente novas é pela mudança do próprio material genético”, isto é, por mutações (página 87). É surpreendente, portanto, vê-lo admitir na sentença seguinte que “tem sido discutido que as mutações poderiam pouco ter a ver com a evolução, porque as que realmente têm sido observadas no laboratório, ou no homem, são quase sempre prejudiciais (ou quando muito, neutras)” (página 87). (Ênfase acrescentada).
“A proporção de uma mutação favorável em mil não parece elevada, porém é provavelmente generosa. … E o total de um milhão de etapas mutacionais parece bastante, porém provavelmente constitui uma subestimativa. … Suponhamos, entretanto, que esses números sejam estimativas razoáveis. Dentro dessa proporção, mas sem seleção alguma, certamente teríamos de cruzar um milhão de linhagens para obter uma contendo duas mutações favoráveis, e assim por diante, até mil elevado à milionésima potência para obter uma linhagem contendo um milhão. Na realidade isso poderia ainda não acontecer, porém esta é uma maneira útil de visualizar as fantásticas probabilidades contrárias à obtenção de numerosas mutações favoráveis em uma linhagem, somente devido ao puro acaso. Ao se escrever o número mil elevado à milionésima potência tem-se o algarismo 1 seguido de três milhões de zeros, o que exigiria três grandes volumes de 500 páginas cada, somente para imprimí-los! … Ninguém apostaria em um acontecimento tão improvável. Não obstante, isso aconteceu! Aconteceu graças à atuação da seleção natural e às propriedades da substância viva que tornam inevitável a seleção natural!” (17) Seleção Natural Logicamente, então, o próximo tópico é a seleção natural. Pode ela realmente converter a impossibilidade de Huxley (e também de Schramm) em algo inevitável? Como Lasker afirma à página 19, antes da época de Darwin foi reconhecida como fato uma espécie contrária de seleção – a seleção normalizadora. “Esse processo, ora denominado de seleção normalizadora, de fato retarda e estabiliza a evolução pela eliminação da descendência que difere grandemente de seus ancestrais”. Entretanto, continua ele a dizer: “Foi o reconhecimento da evolução progressiva, contudo, e sua firme fundamentação nos numerosos fatos estabelecidos por Darwin e seus seguidores, que deu origem a uma revolução no pensamento biológico. A evolução progressiva é a seleção da descendência alterada que seja mais bem adaptada que seus ancestrais” (página 20). Curiosamente, a despeito desses “numerosos fatos estabelecidos por Darwin e seus seguidores”, encontramos George Gaylord Simpson, talvez o mais influente paleontologista evolucionista, admitindo a respeito da seleção natural, em um de seus livros, que “pode ser objetado que a teoria é bastante frágil, e tem condição tão somente de especulação” (18).
“As experiências (com a mariposa) ilustram muito bem a seleção natural em ação ou a sobrevivência do mais apto, porém não mostram a evolução em andamento, pois embora possam as populações alterar seu conteúdo de formas claras, escuras ou intermediárias, todas as mariposas permanecem, do princípio ao fim, as mesmas Biston betularia” (19). Além disso, as mariposas nem mesmo demonstram a seleção natural, que não mais é definida em termos de sobrevivência, mas sim como reprodução diferencial (Glossário, página 382). É duvidosa a evidência quanto a ter havido realmente um aumento na proporção de mariposas escuras que foram reproduzidas. Paul Ehrlich e Richard Holm relatam que em algumas experiências não foi encontrada a deficiência esperada de indivíduos claros. Em outra experiência, os resultados foram inconsistentes – enquanto os descendentes de alguns acasalamentos apresentaram uma proporção significativamente alta de mariposas escuras, os descendentes de outros não apresentaram (20). Ainda mais, em um recente artigo publicado no Scientific American, o autor observa que “as mariposas escuras continuam sendo comuns em áreas onde teoricamente os pássaros as teriam exterminado. Algum fator pode estar contrabalançando a desvantagem das mariposas escuras, talvez atuando durante os estágios de larva ou pupa, que constituem a maior parte da vida das mariposas. Complexidades desse tipo estão ainda para ser investigadas” (21). Parece, portanto, que, excluindo a extinção total de toda a espécie, as mariposas Biston betularia permanecerão polimórficas. Porém, como Lasker ressalta na página 103, a existência de muitos polimorfismos é difícil de entender em termos de seleção natural. De fato, alguns cientistas, incluindo King e Jukes (1969), citados por Lasker, usam os polimorfismos e outras evidências para argumentar que a evolução deve ocorrer mediante variações “neutras”, que não são vantajosas nem desvantajosas, e portanto “invisíveis” à seleção natural. Lasker afirma brandamente: “isto está em desacordo com a teoria da seleção natural” (página 103). Stephen Jay Gould, de Harvard, resumindo esses pontos de vista, é mais categórico: “Todavia, algo ainda mais fundamental está ameaçando o próprio Darwinismo” (22). Parece que as evidências no caso das mariposas apoiam este último ponto de vista, tanto quanto o de que a seleção natural é importante. Darwin estava bem familiarizado com as alterações nas plantas e animais domésticos, resultantes de cruzamentos artificiais, e considerou que fosse exeqüível a extrapolação: “Embora lento o processo de seleção, se tanto pode o débil homem fazer com seu poder de seleção artificial, não posso vislumbrar limite algum à intensidade de alteração, à beleza e à complexidade infinita de coadaptações entre todos os seres orgânicos, uns com os outros, e com as suas condições físicas de vida, que podem ser efetuadas no longo decurso do tempo, pelo poder de seleção da natureza” (23). Contudo, mais de cem anos após Darwin, todas as evidências experimentais indicam que a variação confina-se dentro de limites. Por exemplo, as experiências tão divulgadas relativas às moscas das frutas, com mutações induzidas artificialmente, e seleção artificial, resultaram em considerável variabilidade. Porém tentativas de levar essas variações além de certos limites resultaram em esterilidade em massa, e em morte. Tentativas de manter as variações próximo de seus limites levaram à reversão à norma. Ernst Mayr, eminente evolucionista de Harvard, encara esses resultados como inteiramente normais. “Obviamente qualquer melhoramento drástico obtido pela seleção deve comprometer seriamente o estoque de variabilidade genética. … A resposta mais freqüente da seleção unidirecional é uma diminuição na aptidão geral. Isso afeta virtualmente todas as experiências de cruzamento” (24). Mayr continua a acreditar que grandes alterações são possíveis, porém não cita nenhum caso observado, e concorda que “muito disso obviamente é especulativo” (25). Não somente há uma desconcertante escassez de evidências. Parece também que os evolucionistas não têm sido capazes de surgir com uma definição exeqüível de seleção natural. “Natural” significa “não artificial”, o que até aí está claro. “Seleção”, de acordo com Lasker, significa “que existem variantes biológicas favoráveis em termos das circunstâncias sob as quais sua seleção ocorre” (página 92). À parte o fato de que essa definição pressupõe sua própria conclusão, isto é, que a seleção realmente ocorre, o problema reside em determinar objetivamente quais variantes são favoráveis em termos das circunstâncias de sua ocorrência. Outro eminente evolucionista, G. Ledyard Stebbins, ora na Universidade da Califórnia, em Davis, admitiu: “Obviamente … uma estimativa final da importância da seleção na evolução deve depender grandemente da determinação de quais … as diferenças que são … adaptativas. … Infelizmente, entretanto, a determinação do caráter adaptativo de muitos tipos de diferenças entre organismos é um dos problemas mais difíceis da Biologia” (26). Podemos, portanto, pouco mais fazer do que especular (27). Mayr concorda: “… jamais se pode afirmar com certeza que uma dada estrutura não tenha significado seletivo” (28). Simpson considera isso como objeto de senso comum: “É notória a falibilidade dos juízos pessoais quanto ao valor adaptativo de caracteres particulares, mais particularmente quando ocorrem eles em animais muito distintos dos que hoje vivem” (29). De fato, a maior parte dos evolucionistas concorda hoje que a “aptidão” não pode ser analisada objetivamente. Daí a precaução de Lasker: “A aptidão é definida pela prova da sobrevivência, e não pela ‘aptidão’ conforme algum sentido humano de avaliação” (página 92). Isto significa que os que sobrevivem são mais aptos porque os mais aptos é que sobrevivem. Isto constitui uma tautologia – uma falácia lógica que nada explica (30). Com esse tipo de definição, é fácil ver como a seleção natural pode simultaneamente “explicar” no exemplo de Lasker, a evolução tanto do parto prematuro como do tardio, e mesmo da desmama nos animais (página 93). As deficiências das mutações e da seleção natural como mecanismos da evolução levaram a uma ênfase recente na chamada deriva genética aleatória, acoplada às flutuações nos tamanhos das populações (páginas 107 a 118). Entretanto, como a deriva genética sozinha não pode produzir nada senão a mesma espécie e o mesmo nível de organismo, essa ênfase parece não se justificar. Os interessados poderão ler o breve relato de Mayr sobre o conceito de deriva genética, acompanhado de uma série de aplicações indefensáveis (31). Mayr aparentemente ainda crê que possa haver um papel a ser desempenhado na evolução pela deriva genética. Parece, porém, ter dúvidas sobre o papel de outros mecanismos aleatórios: “Fenômenos aleatórios como a recombinação … introduzem na evolução um considerável grau de indeterminação. Temporariamente eles podem ser ainda mais fortes que a seleção em populações completamente isoladas, e pelo menos inicialmente pequenas. É ainda bastante obscura a importância que apresentam tais populações para a formação de novas espécies, e em última análise para a evolução” (32). Os evolucionistas enfrentam aqui um dilema, pois quanto maior o papel atribuído aos processos aleatórios, menor é a probabilidade de que a impossibilidade de Huxley se converta em uma inevitabilidade pela seleção natural. Creio ter questionado substancialmente as “evidências” de que qualquer mecanismo biológico conhecido possa produzir alterações macroevolutivas. É comumente aceito, entretanto, que a despeito das deficiências dos mecanismos explicativos, o registro fóssil provê claras evidências da macroevolução. Consideram-se, então, os fósseis, a seguir. Hiatos no Registro Fóssil Lasker afirma: “Existem hoje muitas evidências nos fósseis a favor da existência de todas as espécies de formas intermediárias que indicam a natureza do relacionamento entre o homem e outros animais, e novas descobertas continuamente são acrescidas. … Nosso problema não reside propriamente na descoberta de elos perdidos” (página 18). Realmente, o problema inicialmente é semântico. Como Lasker posteriormente declara: “Não se pode propriamente falar de elos perdidos e elos conhecidos na evolução, pois a evolução progride em linhas e não em cadeias, e é sempre possível aprimorar o nosso conhecimento à medida em que se tornam conhecidos mais pontos da linha, e se tornam menores os segmentos desconhecidos existentes entre eles” (página 242). Em outras palavras, ele está afirmando que, como a evolução ocorre de forma contínua, com inumeráveis pontos, não se pode esperar encontrar todas as formas intermediárias sobre a mesma linha. De fato, desde que se pressuponha um continuo evolutivo, a distância entre supostas formas ancestrais intermediárias não apresenta, como afirma Lasker, nenhum “problema”. Ninguém jamais exigiu que se conseguissem todos os fósseis de todos os indivíduos em uma suposta linha de descendência. Porém parece ser razoável exigir que se consiga uma série na qual cada um não difira muito de seu vizinho. Qualquer que seja a suposição, o fato é que os hiatos existentes no registro fóssil entre formas pretensamente relacionadas, são grandes e sistemáticos. Simpson declarou que “é uma característica do registro fóssil conhecido, o aparecimento abrupto da maior parte dos taxa …. Hiatos entre espécies são esporádicos e freqüentemente pequenos . Hiatos entre ordens, classes e fila são sistemáticos e quase sempre grandes” (33). Outro paleontologista, T. Neville George, afirma: “Não mais há necessidade de se excusar pela pobreza do registro fóssil. Sob alguns aspectos ele se tornou quase incontrolavelmente rico, e as descobertas sobrepujam a integração. … O registro fóssil continua sendo composto principalmente de hiatos” (34). Em recente artigo publicado na revista “Evolution”, um paleontologista da Universidade de Oklahoma disse: “Apesar da auspiciosa promessa de que a paleontologia provê uma maneira de ver a evolução, ela tem levantado algumas dificuldades desagradáveis para os evolucionistas, das quais a mais notória é a presença de “hiatos” no registro fóssil . A evolução exige formas intermediárias entre as espécies, e a paleontologia não as provê” (35). (Ênfase acrescentada). Tão grandes são os hiatos, que os supostos relacionamentos ancestrais são altamente arbitrários. Mayr, que sem dúvida é um dos principais taxonomistas americanos, declara: “A maior parte dos ‘taxa’ acima do nível de família são claramente delimitados. Moluscos, pingüins, vespas, e na realidade a maior parte dos ‘taxa’, são separados de seus parentes mais próximos por um indiscutível hiato, muito mais do que acontece na maioria dos gêneros e famílias. Entretanto, permanece certo que as categorias superiores nas quais colocamos esses ‘taxa’ são mal definidas. … Nenhuma norma até agora foi encontrada para a classificação não arbitrária dos ‘taxa’. … É na arbitrariedade da definição que todas as categorias superiores diferem da categoria da espécie” (36). Uma revisão crítica do livro de Mayr declara que “praticamente todos os biologistas devem concordar que a espécie é a única categoria taxonômica que tem uma existência completamente objetiva, pelo menos nos exemplos mais favoráveis” (37).
Há um grupo de fósseis, contudo, a que repetidamente se refere sempre que surge a questão dos hiatos – é a série do cavalo. Ela aparece virtualmente em todos os livros que tratam do assunto da evolução. Não obstante, de conformidade com Simpson “esse exemplo usual tem sido grandemente deturpado” (38). Garrett Hardin ressalta que uma exposição antiga do Museu Americano de História Natural visando indicar a linha evolutiva direta do cavalo foi amplamente fotografada e divulgada antes que sua natureza incorreta fosse esclarecida. Essas fotografias tendenciosas permanecem ainda em alguns livros didáticos (39). Lasker apresenta os fósseis corretamente em um diagrama à página 207. Observe-se que, apesar do “abundante material fóssil”, somente três fósseis são considerados como ancestrais do cavalo moderno, o Equus. São eles o Eohippus, o Miohippus, e o Merychippus. Simpson, em quem Lasker se baseia para construir seu diagrama, é provavelmente a maior autoridade na série do cavalo. Considerem-se as afirmações seguintes do próprio Simpson: “Cada aspecto da evolução do cavalo descreve uma história relativamente complexa ao ser examinada em detalhe e em todas as linhas divergentes da família do cavalo. As patas … dificilmente teriam iniciado sua evolução no Eoceno, e então rapidamente evoluíram em direção a um tipo básico oligoceno de três dedos, que permaneceu praticamente estático em algumas linhas posteriores … e somente em uma linha finalmente evoluíram rapidamente em uma fase em direção a um tipo monodátilo. Isso novamente não continuou a tendência usual entre os tipos tridátilos, mas constituiu uma nova direção evolutiva. Na linhagem específica do Eohippus ao Equus, a mecânica geral dos pés tornou-se inicialmente mais complexa, e depois mais simples. O número de dedos não se alterou continuamente de quatro (na pata dianteira) até um, mas sofreu alteração em duas etapas primeiramente de quatro para três, e depois, muito mais tarde, de três até um, cada transição rápida sendo seguida de ajustamentos mecânicos mais lentos, necessários ao novo tipo de pata, e às alterações no peso dos animais. O cavalo caracteriza ainda a exceção à regra de que os animais tendem a crescer de tamanho em sua evolução. De fato, os cavalos de que se tem notícia no Eoceno recente são em média menores do que o Eohippus do Eoceno. Antes ainda, no Mioceno e no Plioceno, havia pelo menos três ramos distintos da família do cavalo caracterizados pela diminuição do tamanho … enquanto que, ao mesmo tempo, havia outros, de acordo com a “regra”, aumentando de tamanho. Na mesma época, também outros havia flutuando em torno de um tamanho médio, sem alteração apreciável, e ainda outros desenvolviam espécies diferentes, de tamanhos decididamente diferentes como de fato é o caso do Equus hoje” (40). (Ênfases acrescentadas). Dado este tipo de evidências, basta acrescentar a própria declaração de Simpson: “Pode-se confirmar qualquer ‘regra’ que se deseje, desde que se formule a regra e em seguida se interpretem as evidências de conformidade com ela.” (41). Como o diagrama de Lasker indica (Simpson), o prosseguimento apenas dos três estágios presumíveis exige um salto do Velho Mundo para a América do Sul. Além disso, é altamente arbitrária a colocação do Eohippus (mais amplamente chamado de Hyracotherium) como o primeiro elo da série. Em um artigo anterior disse Simpson: “Matthew mostrou e insistiu que o Hyracotherium (incluindo o Eohippus) é tão primitivo (42) que não é muito mais definidamente eqüídeo do que taprídeo, rinocerotídeo, etc., embora seja costumeiro colocá-lo na raiz do grupo eqüídeo” (43). (Ênfase acrescentada). O resto do registro fóssil oferece pouca tranqüilidade aos macroevolucionistas. Simpson afirma que existe um considerável hiato entre o Eohippus (Hyracotherium) e sua suposta ordem ancestral. Continua ele dizendo: “Isto é verdade para todas as trinta e duas ordens de mamíferos. … Os membros mais antigos e mais primitivos de todas as ordens já possuem os caracteres ordinais básicos, e em nenhum caso existe uma seqüência aproximadamente contínua desde uma ordem até outra conhecida. Na maior parte dos casos a descontinuidade é tão distinta e o hiato tão grande que a origem da ordem é especulativa e muito discutida” (44). (Ênfase acrescentada). “Existe … uma tendência em direção a uma deficiência sistemática no registro da história da vida. É portanto possível alegar que tais transições não estão registradas porque não existiram, que as alterações não se deram por transição, mas por saltos bruscos na evolução” (46). (Ênfase acrescentada). Alguns evolucionistas têm aceito os hiatos no registro fóssil tal qual se apresentam, e têm declarado que as grandes categorias devem ter surgido por “macrogênese”, ou mutação sistêmica, ao invés de milhões de pequenos passos (“gradualismo filetico”) conforme proposto por virtualmente todos os outros (47). O mais proeminente daqueles é Richard Goldschmidt, da Universidade da Califórnia, em Berkeley, que usou a expressão “monstro auspicioso”. O livro de Goldschmidt contém uma valiosa crítica de outras idéias evolucionistas (48). Falando do conceito de Goldschmidt, entretanto, Mayr declara: “Está bem comprovada a ocorrência de monstruosidades genéticas por mutação, como por exemplo os mutantes homeóticos na Drosophila (mosca das frutas), porém elas constituem anomalias tão evidentes que esses monstros somente podem ser designados como “desesperançados”. Eles são tão completamente desproporcionados que não teriam a mínima probabilidade de escapar da eliminação mediante seleção estabilizadora. … Crer que tal mutação drástica produziria um novo tipo viável, capaz de ocupar uma nova zona adaptiva, é equivalente a crer em milagres” (49). Simpson também aborda este ponto de vista, que depende de mutações múltiplas, simultâneas: “A probabilidade de mutações múltiplas, simultâneas, parece ser ainda menor, de fato desprezível. A postulação de uma taxa de mutação de 0,00001, e da duplicação da probabilidade de cada mutação por outra mutação no mesmo núcleo, corresponderia às circunstâncias mais favoráveis garantidas pelas evidências experimentais. Com essas postulações, a probabilidade de cinco mutações no mesmo núcleo seria de aproximadamente 10-22. Com uma população média efetiva de cem milhões de indivíduos férteis, e com a duração média de um dia para cada geração, novamente postulações extremamente favoráveis, tal evento seria esperado somente uma vez em cerca de 274 bilhões de anos, ou seja, cerca de cem vezes a idade suposta da Terra. Obviamente, a menos que haja um fator desconhecido que aumente tremendamente a probabilidade das mutações simultâneas, tal processo não teve qualquer parte na evolução” (50). (Ênfase acrescentada). Posição Estratigráfica Um argumento usado pelos macroevolucionistas para evitar o confronto com os hiatos no registro fóssil é a asserção de que as camadas da coluna geológica pelo menos demonstram uma ampla progressão das formas de vida simples para as complexas. Entretanto, o registro fóssil está pleno de inconsistências a esse respeito. Pode-se citar Simpson mais uma vez: “A maior parte (dos phyla) … inicia-se no Cambriano … (até recentemente considerado como o estrato no qual primeiramente evoluiu a vida). Há pouca ordem lógica na seqüência do aparecimento. Os Artrópoda aparecem no registro tão cedo quanto sem dúvida os Protozoa, embora no consenso geral os Protozoa sejam o phylum mais primitivo e os Artrópoda o mais “avançado”, isto é, estruturalmente mais complicado entre os não-cordata (ou Invertebrados, como todos os outros phyla, excetuando-se os Cordata, são freqüentemente chamados). Corais e Briozoários não surgem até o Ordoviciano, embora sejam menos altamente organizados do que muitos grupos que ocorrem no primitivo Cambriano. … (O phylum Cordata) deve ser considerado em algum sentido como o mais progredido de todos os phyla, e é de fato o último a aparecer no registro, embora não muito depois dos Briozoários inferiores” (51).
“É nossa opinião que o espectro dos estados de caráter de derivação primitiva, ou polaridade, deve ser elaborado (e de fato usualmente o é) pelo menos inicialmente, com base em critérios morfológicos (em vez de critérios baseados em seqüência de tempo). A razão desta conclusão é que a seqüência nas rochas pode não oferecer um quadro real da polaridade. A primitividade e a aparente antigüidade não são necessariamente correlacionadas” (52). (Ênfase acrescentada). Esses mesmos autores ressaltam que um dos problemas relacionados com a validade dos dados estratigráficos é que a questão da seqüência é freqüentemente decidida com base nos fósseis; portanto, a argumentação constitui um círculo vicioso: “A principal dificuldade no uso de supostas seqüências ancestral-descendentes para expressar a filogenia é que os dados bioestratigráficos são freqüentemente usados em conjunto com a morfologia na avaliação inicial dos relacionamentos, o que leva a um ciclo vicioso óbvio” (54). Porém, pelo menos tão danosa à idéia de que as seqüências fósseis, tais como a do cavalo, demonstram a evolução progressiva, é a afirmação de que “Todos os organismos são fadados a ser relativamente primitivos em alguns aspectos, e relativamente derivados em outros (“O conceito da evolução mosaica”: De Beer, 1954). É pequena a probabilidade de achar um taxon fóssil que seja primitivo em todos os aspectos, com relação a outro taxon mais recente” (55). De fato, Schaeffer e seus colegas dizem que se fosse para considerar somente a “primitividade” relativa dos fósseis do cavalo, a seqüência poderia tão facilmente ser Equus ® Hyracotherium como Hyracotherium ® Equus(56). Supostos Ancestrais da Humanidade Considerem-se, finalmente, os ancestrais fósseis da humanidade. Lasker segue o pensamento corrente e localiza os supostos ancestrais evolutivos do homem em três categorias: (1) Australopithecus, (2) Homo habilis, e (3) Homo erectus. A última categoria inclui os fósseis do famoso “homem de Pequim” e do “homem de Java”, bem como numerosos outros (página 263). Não se inclui o Homem de Neanderthal, que virtualmente todos os evolucionistas agora incluem no Homo sapiens (página 292). Exatamente quando o livro de Lasker estava sendo escrito em 1973, entretanto, Richard Leakey e outros descobriram novos fósseis na África, que virtualmente eliminaram a primeira categoria, os Australopithecus, como ancestrais do homem. Nas próprias palavras de Leakey, o Crânio 1470 “deixa em ruínas a noção de que todos os fósseis primitivos podem ser arranjados em uma seqüência ordenada de alteração evolutiva” (57). Leakey agora considera o Australopithecus como “simiesco” e “provavelmente mais um parente do que um progenitor da humanidade” (58).
Lasker discute o crânio 1470 nas páginas 264 a 267, baseando suas observações nos primeiros relatórios de Leakey, e afirma que é possível concluir que o 1470 é um “homem verdadeiro”. “Em qualquer caso, os que acreditam ter havido uma longa história do homem verdadeiro têm um novo candidato fóssil para considerar…” (página 265). Ralph L. Holloway Jr., antropologista da Universidade de Columbia, concluiu que o 1470 “era capaz de alguma espécie de linguagem humana” porque o crânio tem “uma saliência conhecida como área de Broca, que não existe nos símios, mas encontra-se no homem, e é considerada como o centro da linguagem” (61). Apesar de sua configuração moderna, Leakey interpreta o Crânio 1470 como um elo ancestral do homem devido à sua pequena capacidade craniana. Contudo Stephen Molnar, editor associado do “American Journal of Physical Anthropology”, depois de comentar a respeito da “utilidade” de comparar capacidades cranianas nos fósseis, fez os seguintes surpreendentes comentários: Nas populações modernas … há um tão amplo intervalo de variação que a extremidade inferior do intervalo situa-se bem abaixo da capacidade de certos hominídeos fósseis, embora não existam evidências de que esses indivíduos sejam menos inteligentes do que pessoas com maiores volumes cranianos. É improvável que as diferenças de tamanho do cérebro nas populações modernas tenham qualquer relevância na variação da capacidade mental – um fator que torna um exercício fútil e sem significado as comparações da capacidade craniana entre grupos modernos. Como afirmou o famoso neuroanatomista von Bonin (1961), a correlação entre o tamanho do cérebro e a capacidade mental não é significativa no homem moderno. Um bom exemplo pode ser visto no caso das mulheres, que têm em média capacidade craniana dez por cento menor do que os homens. Ninguém ousou sugerir que isso indique menor capacidade mental. Muitos homens famosos na história tiveram também, após sua morte, os seus cérebros pesados e medidos. Os valores flutuaram no intervalo do H. sapiens, desde Anatole France (capacidade craniana de aproximadamente 1100 cc) até Oliver Cromwell e Lord Byron (capacidade craniana de aproximadamente 2200 cc). … Variações de mais ou menos 400 cc em torno da média são encontradas na maioria das populações européias. Os indivíduos com capacidades cranianas maiores ou menores são pessoas intelectualmente competentes e de comportamento normal. De fato, existem muitas pessoas com 700 a 800 centímetros cúbicos” 62). (Ênfase acrescentada). Se o tamanho do cérebro nada significa para as populações modernas, não seria justo sugerir que também nada significa para as populações fósseis? Para todos os propósitos práticos o Crânio 1470 de Leakey elimina qualquer imaginário ancestral evolutivo da humanidade. À parte a questão mais ampla, outros antropologistas têm concordado com a eliminação do Australopithecus feita por Leakey. David Pilbeam e Stephen Jay Gould, dois proeminentes especialistas, utilizando técnicas alométricas (63), e Charles F. Oxnard, anatomista e antropologista da Universidade de Chicago, utilizando técnicas de análise multivariacional (64), concluíram que o Australopithecus não foi ancestral do homem. Assim, o Australopithecus está rapidamente a caminho de ser deserdado da ancestralidade humana. A categoria do Homo habilis foi primeiro proposta em 1961 pelo extinto Louis S. B. Leakey (pai de Richard Leakey). Como indica Lasker, o status separado daquela categoria foi posto em dúvida desde o princípio: “Alguns estudiosos do problema acreditam que o grau de variação de tamanho de um grupo de dentes a outro (ou em um dente particular, de uma dimensão a outra) verificado entre o Australopithecus e o Homo habilis ocorre somente entre espécies. Outros (ver por exemplo Brace et al. 1971) chamam a atenção para o fato de que o tamanho dos dentes pode variar grandemente dentro de uma espécie” (página 264). Recentemente Brace e Wolpoff, ambos antropologistas da Universidade de Michigan, fizeram os seguintes comentários em resposta ao artigo de David Pilbeam e Stephen Jay Gould, citado na referência anterior (63) : “A própria existência do taxon Homo habilis é devida mais a uma ampla divulgação jornalística … do que a dados reais … Pilbeam concorda agora conosco, acreditando que a variação dentária nos Australopithecus é tão grande que as mandíbulas e os dentes sozinhos não são de utilidade para distinguir os taxa, conclusão essa recentemente enfatizada pela descoberta de um crânio no lago Rudolph com capacidade de 500 cc e dentes bastante pequenos (ER 1813)”. “A amostra que Pilbeam agora deseja atribuir ao Homo habilis consiste de somente quatro espécimes. … Não há espécime algum simultaneamente com capacidade craniana conhecida e tamanho de dente conhecido. … Não existe um único crânio do Pleistoceno inferior com capacidade craniana conhecida, que esteja associado com qualquer material pós-craniano útil para a determinação do tamanho do corpo. … Isso é verdade também para o Homo erectus. … O variado uso do taxon criou nada mais além de confusão. Portanto reiteramos a sugestão anterior de que o ‘Homo habilis seja formalmente enterrado” (65). Isso deixa somente o Homo erectus para preencher o hiato entre os seres humanos e os não-humanos. Todavia, não há evidências de macroevolução aqui, pois considere-se a declaração de Lasker: “Somente um ou dois desses fósseis caem fora do intervalo de uma pequena série de indivíduos brancos americanos” (página 284). Dado o intervalo de variação entre a população mundial, pode ser dito que o Homo erectus tem qualquer significação evolutiva? S. M. Garn (em quem se baseia Lasker) “considera significativo o aumento do tamanho do crânio do Homo sapiens” (página 284), porém, como foi visto, tais diferenças parecem não significativas, particularmente à luz do fato ressaltado acima por Brace e Wolpoff, de que ignoramos o tamanho do corpo de todos os crânios do Homo erectus. Se a capacidade craniana é significativa, como levar em conta o Homem de Neanderthal? Explicando a grande capacidade craniana de alguns espécimes Neandertais (1,75 litros, em contraposição ao intervalo de 1,30 a 1,45 litros para a média do europeu moderno), Lasker acha necessário acautelar que “não é garantido supor que o homem de Neanderthal fosse em geral significativamente mais inteligente do que o homem subseqüente”. (Página 291). ConclusãoOs argumentos a favor da macroevolução falham em todos os níveis significativos ao serem confrontados com os fatos. A origem da vida, as mutações, a seleção natural, e o registro fóssil, todos falham no apoio à doutrina. Por que então os evolucionistas continuam a asseverar que a evolução é um fato? A resposta é que a evolução foi definida por alguns autores de tal maneira que ela se tornou um fato. Lasker dá uma definição comum da evolução: “Uma alteração contínua ao longo de gerações, nas freqüências das características determinadas geneticamente” (página 376). Assim, pode-se dizer que qualquer alteração na freqüência dos genes, por exemplo “entre pai e filho”, é evolução. Lasker diz que a evolução “é uma alteração lícita na composição genética dos membros de uma população” (página 16, ênfase acrescentada). Ao assim proceder, ele e outros evolucionistas incluem as leis da hereditariedade dentro da definição da evolução. Entretanto isso só poderá levar à confusão, pois como diz Theodosius Dobzhansky: “Todos os seres vivos crescem e reproduzem-se à sua semelhança. … Este processo de auto produção, ou de igual gerar a igual, é a essência da hereditariedade. A hereditariedade é … a antítese da evolução. … A evolução é um processo que fez os descendentes diferentes de seus ancestrais” (66). (Ênfase acrescentada). Essa espécie de não-definição deveria ser deixada de lado. Apesar da “síntese neo-Darwinista”, a ciência da Genética tornou desnecessário explicar a variabilidade em termos evolucionistas. Considere-se a seguinte afirmação de Gould: “… há muito mais variabilidade genética dentro de populações naturais do que a escola clássica poderia possivelmente permitir, e mesmo mais do que muitos geneticistas de campo jamais ousaram imaginar. Em diversos organismos, mais da metade dos genes pesquisados existem em alelos alternados dentro de uma população, ao mesmo tempo em que os indivíduos podem ser variáveis em até 25% de seus genes (embora 5 a 15% por indivíduo seja o intervalo usual)” (67). Assim, o que tem sido considerado como evidências de alterações evolutivas, pode ser explicado simplesmente como variação genética normal. E, como tentei mostrar, não há evidências, tanto no registro fóssil e nas observações da natureza, como na experimentação biológica, de que essas variações podem estender-se através das limitações genéticas naturais, e produzir alterações macroevolutivas. Agradecimentos As seguintes pessoas leram trechos deste artigo, durante sua preparação. Embora partilhem das dúvidas do autor a respeito da macroevolução, não são elas responsáveis, de modo algum, por quaisquer afirmações particulares feitas no artigo. Phillip Coleman, Professor de Biologia no Sacramento City College. Bibliografia Geral Sou devedor às seguintes fontes por muitas das idéias expressas neste artigo. São elas recomendadas para quem desejar aprofundar-se no assunto: Gish, Duane T. 1972. Speculations and experiments related to theories on the origin of life: a critique. ICR Technical Monograph nº 1. Creation Life Publishers, San Diego, California. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Artigo publicado naFolha Criacionista 16 |
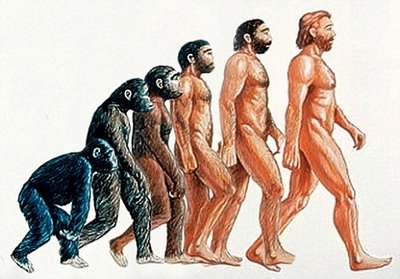
A MACROEVOLUÇÃO QUESTIONADA
Artigos
Nenhum artigo encontrado.
Notícias
Nenhuma notícia encontrada.
 Além do mais, esses compostos são destruídos também pelo Oxigênio. Assim, os defensores da geração espontânea da vida são levados a admitir que a atmosfera primitiva não continha Oxigênio (Ver a figura III.3, página 26, do livro de Lasker). Atualmente, a atmosfera contém cerca de 21% de Oxigênio.
Além do mais, esses compostos são destruídos também pelo Oxigênio. Assim, os defensores da geração espontânea da vida são levados a admitir que a atmosfera primitiva não continha Oxigênio (Ver a figura III.3, página 26, do livro de Lasker). Atualmente, a atmosfera contém cerca de 21% de Oxigênio. As mutações são fundamentais para a evolução, não obstante os fatos observados novamente pareçam contradizer a doutrina. Lasker livra-se do problema afirmando que “ninguém alegaria que a mutação sozinha produz evolução. De fato, ela produziria a evolução na presença de outros fatores, como por exemplo a seleção natural …” (página 87). Considerem-se, porém, os seguintes cálculos feitos por Sir Julian Huxley com relação à suposta evolução do cavalo (e então multiplique-se o resultado pelo número dos supostos passos evolutivos entre a ameba e o homem):
As mutações são fundamentais para a evolução, não obstante os fatos observados novamente pareçam contradizer a doutrina. Lasker livra-se do problema afirmando que “ninguém alegaria que a mutação sozinha produz evolução. De fato, ela produziria a evolução na presença de outros fatores, como por exemplo a seleção natural …” (página 87). Considerem-se, porém, os seguintes cálculos feitos por Sir Julian Huxley com relação à suposta evolução do cavalo (e então multiplique-se o resultado pelo número dos supostos passos evolutivos entre a ameba e o homem): Antes que eu seja acusado de citar Simpson fora do contexto, apresso-me a aduzir que o problema de Simpson não é a comprovação de que a seleção ocorra, porém a demonstração de que ela tenha qualquer efeito na evolução. O Professor Lasker apresenta o exemplo favorito dos evolucionistas: a mariposa
Antes que eu seja acusado de citar Simpson fora do contexto, apresso-me a aduzir que o problema de Simpson não é a comprovação de que a seleção ocorra, porém a demonstração de que ela tenha qualquer efeito na evolução. O Professor Lasker apresenta o exemplo favorito dos evolucionistas: a mariposa 
 É instrutiva, neste aspecto, a discussão apresentada por Lasker sobre
É instrutiva, neste aspecto, a discussão apresentada por Lasker sobre  O registro fóssil é tão equívoco, de fato, que três proeminentes paleontologistas concluíram recentemente que a posição estratigráfica nem mesmo devia ser considerada na determinação inicial da filogenia (linha da suposta descendência evolutiva):
O registro fóssil é tão equívoco, de fato, que três proeminentes paleontologistas concluíram recentemente que a posição estratigráfica nem mesmo devia ser considerada na determinação inicial da filogenia (linha da suposta descendência evolutiva): A razão para as dramáticas afirmações de Leakey é que o Crânio 1470, exceto pelo seu pequeno tamanho, cerca de 800 cc, é aparentemente de forma mais “moderna” do que quaisquer dos até aqui supostos ancestrais do homem. Por exemplo, a fronte inclinada e os seios frontais proeminentes dos fósseis do Homo erectus não estão presentes no Crânio 1470 (59). Não obstante, o Crânio 1470 é considerado como tendo cerca de 1,8 milhões de anos – um contemporâneo do Australopithecus, e mais velho do que qualquer fóssil do Homo erectus (60).
A razão para as dramáticas afirmações de Leakey é que o Crânio 1470, exceto pelo seu pequeno tamanho, cerca de 800 cc, é aparentemente de forma mais “moderna” do que quaisquer dos até aqui supostos ancestrais do homem. Por exemplo, a fronte inclinada e os seios frontais proeminentes dos fósseis do Homo erectus não estão presentes no Crânio 1470 (59). Não obstante, o Crânio 1470 é considerado como tendo cerca de 1,8 milhões de anos – um contemporâneo do Australopithecus, e mais velho do que qualquer fóssil do Homo erectus (60).